Poznatky, zķskané pųi studiu pųedchozķch kapitol, popisujķcķch pųedevšķm pųķklady z technické praxe, młžeme využķt v ųadģ dalšķch disciplķn.
Ukažme si to alespoņ na jedné oblasti, kterį se v poslednķ dobģ značnģ rozvķjķ, a to na simulaci ekologickżch systémł
(ekosystémł).
V této kapitole budou postupnģ pųedstaveny matematické modely rłznżch ekologickżch systémł a simulace jejich chovįnķ na čķslicovém počķtači
[1, 9, 12]. Nejjednoduššķm modelem, kterż uvedeme, je simulace chovįnķ vżšky hladiny ledovcového jezera v zįvislosti na pųķtoku
vody. Na pųķkladu vģnovaném rłstu populace baktériķ ukįžeme postup pųi nįvrhu nejjednoduššķho modelu systému a jeho nįsledné zdokonalovįnķ. Pųitom
pųedpoklįdįme, že baktérie majķ neomezenż pųķsun živin. Systém tedy nebude obsahovat žįdnou vstupnķ veličinu, vżstupnķ veličinou bude počet baktériķ
v populaci, označķme ji y(t).
Vyšetųujeme ledovcové jezero bez pųirozeného odtoku. Definovali jsme systém, jehož vstupnķ veličina u(t) je pųķtok vody do jezera [m3s-1] a vżstupnķ veličina y(t) je vżška hladiny v jezeųe [m]. Provedeme identifikaci zįvislosti vżšky hladiny jezera na pųķtoku. Pųi identifikaci provedeme nģkolik zjednodušenķ. Vždy se budeme snažit postihnout hlavnķ vlivy na chovįnķ systému a potlačit vlivy podružné. Tķm značnģ zjednodušķme postup prįce.
Matematickż model ledovcového jezera
Pųi stanovenķ zįkonitosti v chovįnķ jezera využijeme platnosti zįkona o zachovįnķ hmoty. Za krįtkż časovż śsek (označķme ho
![]() t) pųiteče do jezera množstvķ vody rovné
t) pųiteče do jezera množstvķ vody rovné ![]() t.u(t) (pro zjednodušenķ pųedpoklįdįme, že pųķtok vody bģhem okamžiku
t.u(t) (pro zjednodušenķ pųedpoklįdįme, že pųķtok vody bģhem okamžiku ![]() t je konstantnķ). Za stejnż okamžik se z jezera odpaųķ čįs vody. Množstvķ odpaųené vody zįvisķ na rychlosti odpaųovįnķ k1[ms-1] a ploše hladiny S [m2].
Celkovģ se za okamžik
t je konstantnķ). Za stejnż okamžik se z jezera odpaųķ čįs vody. Množstvķ odpaųené vody zįvisķ na rychlosti odpaųovįnķ k1[ms-1] a ploše hladiny S [m2].
Celkovģ se za okamžik ![]() t odpaųķ množstvķ vody rovné
t odpaųķ množstvķ vody rovné ![]() tk1S. Zbytek vody, kterż se neodpaųķ, se v jezeųe uložķ a zpłsobķ zvżšenķ hladiny o vżšku
tk1S. Zbytek vody, kterż se neodpaųķ, se v jezeųe uložķ a zpłsobķ zvżšenķ hladiny o vżšku
![]() y (pųitom pųedpoklįdįme, že plocha hladiny je pro celż pųķrłstek vżšky
y (pųitom pųedpoklįdįme, že plocha hladiny je pro celż pųķrłstek vżšky
![]() y konstantnķ), takže se v jezeųe uložķ
množstvķ vody rovné
y konstantnķ), takže se v jezeųe uložķ
množstvķ vody rovné ![]() yS. Celį rovnice zįkona o zachovįnķ hmoty je pak :
yS. Celį rovnice zįkona o zachovįnķ hmoty je pak :
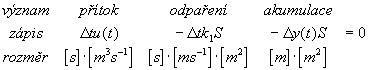
Všimnģme si v rovnici (4.1) provedené rozmģrové kontroly. Rozmģry všech sčķtancł v rovnici musķ bżt stejné. Rozmģrovį
kontrola je nejjednoduššķ zpłsob, jak si ovģųit sprįvnost postupu pųi sestavovįnķ rovnice. Nemusķ však ukįzat všechny naše
chyby. Rovnici (4.1) upravķme dģlenķm ![]() t a
provedeme limitnķ pųechod
t a
provedeme limitnķ pųechod ![]() --> d. Dostaneme diferenciįlnķ rovnici :
--> d. Dostaneme diferenciįlnķ rovnici :
![]()
kterį popisuje chovįnķ vżšky hladiny jezera v zįvislosti na pųķtoku vody. V rovnici vystupujķ parametry k1 a S, které však nejsou konstantnķ. Zatķmco rychlost odpaųovįnķ zįvisķ na okolķ systému (teplota a vlhkost okolķ), plocha hladiny zįvisķ na vżšce hladiny a profilu jezernķho śdolķ. Rychlost odpaųovįnķ młžeme zastoupit jejķ stųednķ hodnotou.
Zįvislost plochy jezera na vżšce hladiny však musķme vyjįdųit, napų. graficky, viz obr. 4.1. Zįvislost plochy jezera na vżšce hladiny je zde popsatelnį nejspķše nģjakou kvadratickou funkcķ. Pro zjednodušenķ modelu provedeme linearizaci zįvislosti plochy hladiny na vżšce hladiny. Skutečnou zįvislost popķšeme pomocķ lineįrnķ zįvislosti, jak je ukįzįno na obr. 4.1. Pųitom musķme zajistit, aby linearizovanż prłbģh prochįzel počįtkem souųadnicového systému, napų. posunutķm počįtku souųadnicového systému. Linearizace je velmi častż zpłsob zjednodušovįnķ modelł. Zįvislost plochy hladiny na vżšce hladiny je pak:
![]()
Z rovnic (4.2) a (4.3) pak młžeme vyjįdųit zįvislost derivace vżšky hladiny na čase (to je vlastnģ rychlost zmģn vżšky hladiny) :
![]()
Tķm jsme zķskali matematickż popis chovįnķ vżšky jezera a současnģ také jeho matematickż model. Všimnģme si však dłležité skutečnosti. Pųi śpravģ rovnice jsme provedli dģlenķ plochou S. To je možné jen tehdy, pokud nemłže bżt plocha hladiny S nulovį, nebo-li pųi prįci s modelem takovż pųķpad musķme vyloučit.
Simulačnķ model ledovcového jezera
Ukažme si nynķ sestavenķ simulačnķho modelu systému vżšky hladiny jezera, kterż byl popsįn rovnicķ (4.4). Nejprve sestavķme bloky, které vytvoųķ hodnotu pravé strany rovnice. Tak zķskįme hodnotu derivace vżšky hladiny. Tuto hodnotu budeme integrovat, čķmž zķskįme hodnotu vżšky hladiny. Celż simulačnķ model je na obr. 4.2 {soubor JEZERO.SIP}. Jako vstupnķ veličina je pųiveden jednotkovż skok polohy (Heavisedełv skok), ųešenķm modelu pro čas t = <0; 200> [den] zķskįme pųechodovou charakteristiku systému, viz obr. 4.3.
Verifikace simulačnķho modelu ledovcového jezera
Jakmile jsme sestavili simulačnķ model chovįnķ jezera, ovģųķme si sprįvnost modelu srovnįnķm s experimentįlnģ namģųenżmi śdaji. Vychįzķme pųitom ze situace, kdy pųķtok vody (vstupnķ veličina) do jezera je stįlż, takže vżška hladiny (vżstupnķ veličina) se ustįlila na konstantnķ hodnotģ, protože pųķtok vody a odpaųovįnķ je v rovnovįze. Současnż pųķtok u(0) = 0,2 m3den-1 a současnou vżšku hladiny y(0) = 2 m označķme jako nulovou, u(0) = 0, y(0) = 0. Nynķ pųivedeme nįhle (skokem) dodatečnż pųķvod vody o jednotku množstvķ za jednotku času (napų. o 0,8 m3den-1). Čas ve kterém se tak stane označķme t = 0. Prłbģh vżšky hladiny (pųechodovį charakteristika systému) je uvedena na obr. 4.4.
Zvyšovįnķ hladiny zpłsobuje také zvģtšenķ plochy hladiny a tķm také zvģtšenķ odpaųovįnķ vody. Proto po určitém čase dojde k ustįlenķ
hladiny na nové hodnotģ. Systém vykazuje setrvačnost vłči zmģnįm vstupnķ veličiny.
Pokud by se jezero nalézalo v śdolķ se svislżmi stģnami, pak by se hladina nezvģtšovala (tedy ani odpaųovįnķ) a hladina jezera by neustįle
stoupala, viz obr. 4.5.
Takovż systém nazżvįmeintegračnķ. Je zajķmavż také tķm, že pro nģj nelze určit statickou charakteristiku, nebo neexistuje ustįlenż stav pro jiné, než nulové hodnoty vstupnķ veličiny.
Nynķ młžeme provést srovnįnķ pųechodové charakteristiky zķskané ųešenķm modelu s namģųenou pųechodovou charakteristikou a verifikovat (zkontrolovat) sprįvnost modelu, pųķpadnģ ho upravit. Tķm je model pųipraven pro experimentovįnķ.
Experimenty s modelem ledovcového jezera
Zįvģrečnou fįzķ a také cķlem celého snaženķ, je provedenķ simulace chovįnķ systému pųi požadovanżch vstupnķch veličinįch. Do simulačnķho
modelu systému pųivedeme vstupnķ veličiny požadovanżch prłbģhł a vyųešenķm zjistķme odezvu vżstupnķch veličin.
Na modelu vżšky hladiny jezera ųešme kolķsįnķ vżšky hladiny v zįvislosti na pųķtoku vody, kterż je popsįn rovnicķ :
u(t) = 0,3 - 0,1cos(0,01t)
K experimentu využijeme model systému vżšky hladiny jezera z obr. 4.2. V čase t = 0 je systém v ustįleném stavu, nebo podle rovnice (4.4) popisujķcķ model systému je derivace vżšky hladiny podle času pųi zadanżch počįtečnķch podmķnkįch nulovį, to znamenį, že vżška hladiny se nemģnķ. Mķsto bloku č. 6 však pųipojķme bloky realizujķcķ požadovanou hodnotu vstupnķ veličiny, viz obr. 4.6 {soubor JEZERO1.SIP}.
Na obr. 4.7 vidķme vżslednou zįvislost vżšky hladiny na hodnotģ vstupnķ veličiny popsané rovnicķ (4.5). Vidķme, že vstupnķ veličina kolķsį mezi hodnotami <0,2; 0,4> [m3den-1] a zpłsobuje zmģnu hladiny v rozsahu <2, 4> [m].
V simulačnķm experimentu budeme dįle pokračovat vyšetųovįnķm vlivu človģka na systém. Napų. se u jezera mį postavit chata, kterį bude odebķrat čįst vody
pro své potųeby. Našķm śkolem je zjistit vliv odbģru vody na chovįnķ hladiny jezera. Odbģr vody človģkem zpłsobķ zmenšenķ pųķtoku vody, pųedpoklįdejme,
že tato situace nastane skokem v čase t = 300 dnķ {soubor JEZERO2.SIP}.
Na obr. 4.8 vidķme chovįnķ hladiny pro dva
rłzné odbģry vody. Odbģr 0,01 m3den-1 zpłsobil témģų nepozorovatelnż rozdķl oproti systému bez odbģru človģkem
(zobrazeno tučnou čarou). Pųi odbģru 0,1 m3den-1 (100 litrł vody za den) dojde k prudkému pųechodu a hladina
dįle kolķsį v hodnotįch o celż metr nižšķch (zobrazeno tenkou čarou). Vliv na systém je tedy podstatnż.
Zįvģr
Po sestavenķ matematického modelu chovįnķ vżšky hladiny ledovcového jezera v zįvislosti na pųķtoku vody jsme zķskali možnost simulačnģ ovģųit vliv zįsahu človģka na tento systém a zķskat tak znalosti potųebné k rozhodnutķ o povolenķ nebo zakįzįnķ tohoto zįsahu. Jak se ukįzalo, byl by požadovanż zįsah pųķliš razantnķ a podstatnģ by ovlivnil vżšku hladiny jezera. Nįslednģ pak młžeme zkoumat jakż vliv by mģla zmģna vżšky hladiny na rostliny a živočichy v jezeųe apod.
Druhżm pųķkladem ( prvnķm je ledovcové jezero ) je ųešenķ rłstu bakteriįlnķ populace. Na tomto pųķkladu si podrobnģji ukįžeme postupné zpųesņovįnķ modelu od nejjednoduššķho a nejménģ pųesného až k modelu velmi dobųe popisujķcķmu chovįnķ bakteriįlnķ populace.
Matematickż model systému exponenciįlnķho rłstu populace
Pro sestavenķ nejjednoduššķho modelu pųedpoklįdįme, že rychlost rłstu baktériķ (derivace) vżstupnķ veličiny podle času) je śmģrnį počtu baktériķ. Pųedpoklįdįme pųitom, že tato śmģrnost je lineįrnķ.
Takto zķskįme diferenciįlnķ rovnici popisujķcķ rłst bakteriįlnķ populace:
![]()
kde je k1 - frekvence dģlenķ baktériķ [hod-1],
y(t) - vżstupnķ veličina - počet baktériķ.
Jak uvidķme na vżsledku simulace rłstu populace, počet jedincł v čase poroste exponenciįlnģ, proto se o rovnici (4.6) hovoųķ jako o rovnici
exponenciįlnķho rłstu.
Simulačnķ model systému exponenciįlnķho rłstu populace
Nynķ sestavķme simulačnķ model systému. Pųedpoklįdįme, že počįtečnķ počet baktériķ je y(0) = 1000 a frekvence dģlenķ baktériķ je k1
= 0,7 hod-1.
Nejprve definujeme použité konstanty :
YNULA = 1000,
K1 = 0,7 .
Nynķ začneme sestavovat vlastnķ simulačnķ model. Zįkladem bude blok INT realizujķcķ numerickou integraci. Na jeho vstup pųivedeme hodnotu derivace vżstupnķ veličiny a na jeho vżstupu dostaneme vżstupnķ veličinu. Jako parametry pųitom zadįvįme pųedevšķm počįtečnķ hodnotu vżstupnķ veličiny YNULA. Vżstupnķ hodnotu z integrįtoru pųivedeme do bloku ZES, kterż realizuje nįsobenķ konstantou, jako konstantu zadįme hodnotu K1 a vżstup z bloku ZES pųivedeme na vstupy integrįtoru. Tķm je celż model definovįn, viz obr. 4.9. {soubor BAKTER1.SIP}.
Nynķ zavedeme simulačnķ model do programu SIPRO. K provedenķ simulace zadįme potųebné časové śdaje. Čas simulace začķnį vždy pro t = 0 hod, zadįme tedy koncovż bod t = 10 hod. Dįle zadįme krok ųešenķ numerické integrace, krok ųešenķ = 0,01. Krok uklįdįnķ určuje které vypočtené body budou uloženy pro zobrazenķ grafu. Omezenķ velikosti pamģti počķtače zpłsobuje, že lze takto uložit nejvżše 1600 vypočtenżch bodł. Zvolķme tedy krok uloženķ = 0,1. Tķm je nastavenķ ukončeno. Spustķme provedenķ vżpočtu a zobrazķme časovż graf ųešenķ, viz obr. 4.10.
Matematickż model systému logistického rłstu populace
Rovnice (4.6) nevystihuje skutečnost, že zįroveņs bunģčnżm dģlenķm probķhį odumķrįnķ starżch bunģk a proto populace ve skutečnosti pųirłstį pomaleji. Po určitém čase dojde k rovnovįze mezi pųķrłstkem a śbytkem baktériķ a počet baktériķ se ustįlķ na konstantnķ hodnotģ (ustįlenż stav). Do matematického modelu proto musķme zahrnout korekci. Pųedpoklįdįme pųitom, že odumķrįnķ baktériķ zpłsobķ zmenšenķ frekvence dģlenķ baktériķ. Toto zmenšenķ bude śmģrné počtu jedincł v populaci. Tak dostaneme tzv. logistickou rovnici rłstu populace :
![]()
kde je k1 - frekvence dģlenķ baktériķ bez odumķrįnķ [hod-1],
k2 - konstanta śmģrnosti odumķrįnķ baktériķ [hod-1].
Simulačnķ model systému logistického rłstu populace
Simulačnķ model logistického rłstu
populace je na obr. 4.11 {soubor BAKTER2.SIP}.
Na obr. 4.12 vidķme vżsledek simulace a zųetelné ustįlenķ počtu baktériķ v populaci na hodnotģ
y(![]() ) = 1 000 000.
) = 1 000 000.
Matematickż model systému rłstu populace podle Hutchinsonovy rovnice
Dalšķm zpųesnģnķm modelu je zahrnutķ skutečnosti, že aktuįlnķ ovlivnģnķ rychlosti množenķ baktériķ nezįvisķ na současném počtu baktériķ, ale na počtu baktériķ v minulé generaci. Zavedenķm této skutečnosti do matematického modelu zķskįme tzv. Hutchinsonovu rovnici rłstu:
![]()
kde je Td - délka bunģčného cyklu [hod].
Simulačnķ model systému rłstu populace podle Hutchinsonovy rovnice
Jejķ simulačnķ model je na obr. 4.13{souborBAKTER3.SIP}.
Na obr. 4.14 vidķme vżsledek simulace a zųetelné ustįlenķ počtu baktériķ v populaci na hodnotģ
y(![]() ) = 1 000 000.
) = 1 000 000.
Zįvģr
Postupnżm zvyšovįnķm složitosti modelu jsme dospģli až do situace, kdy zķskanż matematickż model dosahuje dobré shody se skutečnostķ.
V tomto pųķkladu navįžeme na pųedchozķ pųķklad, ve kterém jsme rozebķrali rłst bakteriįlnķ populace. Vychįzeli jsme pųitom z pųedpokladu, že zdroje rłstu nejsou nijak omezeny. Ve skutečnosti tomu tak nenķ. Proto se nynķ budeme zabżvat vlivem postupného vyčerpįvįnķ zdrojł na rłst populace.
Matematickż model rłstu v prostųedķ s omezenżmi zdroji
Pųi sestavovįnķ matematického modelu vyjdeme z nįsledujķcķch śvah. Množstvķ dostupnżch zdrojł se v zįvislosti na velikosti populace v čase mģnķ. Pųitom platķ:
![]()
kde je z - aktuįlnķ množstvķ dostupnżch zdrojł,
z0 - počįtečnķ množstvķ dostupnżch zdrojł,
k3 - spotųeba zdrojł jednoho jedince [hod-1].
Vzhledem ke skutečnosti, že počet jedincł v populaci nemłže bżt zįpornż, musķ množstvķ dostupnżch zdrojł stįle klesat, až klesnou pod použitelnou mez
(vyčerpajķ se).
Vliv klesajķcķho množstvķ dostupnżch zdrojł se zųejmģ bude projevovat také pųķmo na rłstu populace. Tento vliv bude zųejmģ nepųķmo śmģrnż
množstvķ dostupnżch zdrojł, pųipadajķcķch na jednoho jedince:
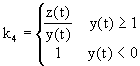
kde je k4 - množstvķ dostupnżch zdrojł pųipadajķcķ na bakterii.
Rłst populace je pak popsįn nįsledujķcķ rovnicķ:
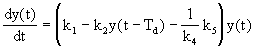
kde je k5 - konstanta śmģrnosti odumķrįnķ baktériķ zpłsobeném nedostatkem dostupnżch zdrojł [hod-1].
Simulačnķ model rłstu v prostųedķ s omezenżmi zdroji
Odpovķdajķcķ simulačnķ model je uveden na obr. 4.15 {soubor BAKTER4.SIP}.
Na obr. 4.16 vidķme prłbģh rłstu populace, kterż je na svém počįtku mįlo ovlivnģn nedostupnostķ zdrojł, proto je prakticky stejnż, jako vżvoj v prostųedķ neomezenżch zdrojł. Postupnżm poklesem množstvķ dostupnżch zdrojł se populace začķnį snižovat, až nįhle velmi rychle zanikį.
Na obr. 4.17 je zkoumįn vliv snķženķ frekvence dģlenķ bakteriķ z hodnoty k1 = 0,7 na k1 = 0,6. Doba života kolonie se prodloužila za cenu zmenšenķ jejķ velikosti, rychlost jejķho zįniku je vżraznģ nižšķ, ale zįnik kolonie je samozųejmģ nevyhnutelnż.
Zįvģr
Simulacķ jsme zjistili jak se bude vyvķjet populace v prostųedķ s omezenżmi zdroji. Zajķmavż je pųedevšķm prudkż zįnik populace, jakmile množstvķ dostupnżch
zdrojł vżraznģ poklesne.
Velmi zajķmavż je systém osķdlovįnķ ostrova jednotlivżmi druhy organismł. Systém je zįvislż pouze na pųedchozķm počtu druhł na ostrovģ ( tķmto se zabżvala pųedchozķ kapitola rłst v prostųedķ s omezenżmi zdroji ). Systém tedy nemį vstupnķ veličinu, vżstupnķ veličinou je počet druhł na ostrovģ.
Matematickż model systému osķdlovįnķ ostrova
Model systému je obvykle sestavovįn nįsledovnģ. Za jednotku času pųibude na ostrovģ počet druhł nepųķmo śmģrnż vzdįlenosti ostrova od pevniny a nepųķmo śmģrnż počtu druhł na ostrovģ. Za stejnou jednotku času vymųe na ostrovģ počet druhł pųķmo śmģrnż na aktuįlnķm počtu druhł. Model je tedy obvykle sestavovįn v nįsledujķcķ podobģ :
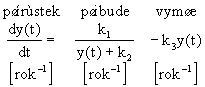
kde je
k1 - konstanta rychlosti pųķrłstku druhł zįvislį na pųevrįcené hodnotģ vzdįlenosti ostrova [rok-1],
k2 - konstanta rychlosti pųķrłstku druhł - v pomģru s konstantou k1 určuje pųķrłstek druhł na neosķdleném ostrovģ,
k3 - konstanta rychlosti vymķrįnķ druhł na ostrovģ [rok-1].
Simulačnķ model systému osķdlovįnķ ostrova
Odpovķdajķcķ simulačnķ model je na obr. 4.18 {soubor OSTROV.SIP}.
Zįvģr
Na obr. 4.19 vidķme vżsledek simulace a zųetelné ustįlenķ počtu na hodnotģ y(![]() ) = 27. Použité hodnoty konstant odpovķdajķ osķdlovįnķ ostrova Krakatoa ptactvem po katastrofģ v roce 1883 do roku 1921.
) = 27. Použité hodnoty konstant odpovķdajķ osķdlovįnķ ostrova Krakatoa ptactvem po katastrofģ v roce 1883 do roku 1921.
Systém pųķjmu potravy ( kterż nepųķmo souvisķ s kapitolou osidlovįnķ ostrova ) je popsįn pomocķ čtyų stavovżch veličin, které definujķ jeho okamžitż stav (nģkdy jsou
označovįny jako kompartmenty). Na obr. 4.20 je zobrazena interakce jednotlivżch prvkł systému (popsanżch
stavovżmi veličinami) za jednotku času. Vżznam stavovżch veličin je nįsledujķcķ :
C1 - koncentrace určité složky potravy v žaludku,
C2 - koncentrace určité složky potravy v krvi,
C3 - koncentrace určité složky potravy ve tkįnķch,
C4 - koncentrace určité složky potravy v moči.
Matematickż model systému pųķjmu potravy
Za jednotku času dojde k pųedįnķ určité složky potravy mezi jednotlivżmi prvky systému a tķm ke zmģnģ hodnot stavovżch veličin, které je popisujķ. Jednotlivé vżmģny za jednotku času popisujķ konstanty uvedené na obr. 4.20. Na jeho zįkladģ młžeme zapsat matematickż model systému :
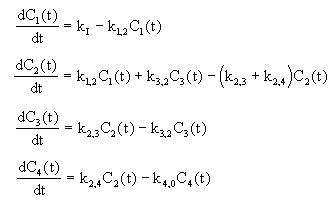
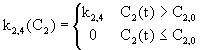
Simulačnķ model systému pųķjmu potravy
Na obr. 4.22 {soubor POTRAVA.SIP} je uveden simulačnķ
model systému pųķjmu potravy pro pųķpad, kdy v čase t = 0 byl žaludek naplnģn potravou
C1(0) = 10 [jednotka potravy] a dalšķ pųķsun potravy se neuskutečņuje (kI= 0). Skutečnost, že pųesun složky potravy
z krve do moče probķhį až po pųekročenķ určité hladiny koncentrace je realizovįna
pomocķ blokł č. 13 a 14. Na obr. 4.23 vidķme vżsledek chovįnķ systému.
Hotovż model využijeme k simulaci chovįnķ systému pųķjmu potravy pųi dįvkovém pųķsunu potravy. Potrava bude pųivįdģna
do systému v malżch množstvķch v pravidelnżch intervalech. Model systému doplnķme o bloky realizujķcķ
tuto skutečnost, viz obr. 4.21 {soubor POTRAVA2.SIP}. Současnģ zmģnķme počįtečnķ podmķnku C1(0) = 0.
Na obr. 4.24 je zobrazen prłbģh vstupnķ veličiny do systému (pųķsun potravy), na obr. 4.25 pak chovįnķ systému.
Zįvģr
Na simulačnķm modelu jsme ukįzali chovįnķ systému pųķjmu potravy. Jednoduchou zmģnou vstupnķho pųķsunu potravy młžeme zkoumat jeho vliv na chovįnķ systému, zįtģž jednotlivżch prvkł systému apod.
Často použķvanżm je model systému koųist - dravec ( což je zjednodušenį aplikace kapitoly systém pųķjmu potravy ), viz obr. 4.26. Pųedpoklįdįme, že koųist mį neomezenż pųķsun potravy a dravci se živķ jen touto koųistķ. Jednį se o dosti podstatné zjednodušenķ, ale jak si ukįžeme i tak młžeme učinit ųadu zajķmavżch zjištģnķ.
Matematickż model systému koųist-dravec
Systém młžeme popsat dvojicķ rovnic. Prvnķ rovnice popisuje zmģnu množstvķ koųisti. Pųedpoklįdįme pųitom, že zmģna množstvķ koųisti je zpłsobena pųķrłstkem koųisti dķky rozmnožovįnķ a hubenķm koųisti dravci, pųirozenį śmrtnost koųisti se neuvažuje:
kde je
x(t) - množstvķ koųisti,
y(t) - množstvķ dravcł,
k1 - konstanta určujķcķ relativnķ rodivost koųisti za jednotku času,
k2 - konstanta určujķcķ relativnķ koųistģnķ (je zįvislé jak na množstvķ dravcł, tak množstvķ koųisti).
Druhį rovnice popisuje zmģnu množstvķ dravcł za jednotku času :
kde je k3 - parametr śčinnosti pųemģny energie z biomasy koųisti na biomasu dravcł,
k4 - relativnķ śmrtnost dravcł.
Simulačnķ model systému koųist-dravec
Na obr. 4.27 {soubor KOR_DRAV.SIP} je uveden simulačnķ model systému koųist-dravec. Na obr. 4.28 vidķme vżsledek chovįnķ systému pro dva pųķpady. Tučnou čarou je zobrazeno chovįnķ systému bez zįsahu človģka. Pųedpoklįdanį rodivost koųisti je k1 = 0,4. Tenkou čarou je zobrazen systém se zįsahem človģka, kterż odlovķ čįst koųisti, toto odlovenķ je śmģrné aktuįlnķmu množstvķ koųisti a proto ho młžeme do modelu zavést jako snķženķ rodivosti koųisti na hodnotu k1 = 0,2. Vidķme, že prłmģrnį hodnota, kolem které kmitalo množstvķ koųisti złstala zachovįna, pouze se zmenšila amplituda kmitįnķ. U dravcł však dojde ke snķženķ prłmģrného množstvķ dravcł. Znamenį to, že zįsah ve skutečnosti ovlivnil hlavnģ dravce.
Na stejném modelu nynķ ukažme vliv človģka, kterż zpłsobķ svżmi zįsahy zvżšenķ relativnķ śmrtnosti dravcł o 50%, z hodnoty k4 = 1,2 na hodnotu k4 = 1,8. Na obr. 4.30 je tučnģ zobrazeno chovįnķ systému bez zįsahu človģka, tence se zįsahem. Vidķme, že prłmģrnį hodnota, kolem které kmitalo množstvķ koųisti se vżraznģ zvżšila, amplituda kmitįnķ se zvżšila rovnģž. Množstvķ dravcł sice sleduje vżvoj množstvķ koųisti, ale v minimech je množstvķ dravcł tak malé, že hrozķ jejich śplné vyhubenķ.
Verifikace simulačnķho modelu systému koųist-dravec
Provedené simulačnķ vżstupy je nutno nynķ verifikovat s vżsledky pozorovįnķ systému v pųķrodģ, młže to bżt napų. systém hraboši-kįņata, viz obr. 4.32. Pųitom si musķme uvģdomit, že již tato pozorovįnķ jsou z technického pohledu zatķžena dosti značnou chybou a to ųįdovģ i pųes 5 - 10%. Dosaženķ shody simulačnķho vżstupu s pozorovįnķm s pųesnostķ na 5 - 10% je tedy možno považovat za dostatečné.
Experimenty s modelem systému koųist-dravec
Simulačnķ model je nynķ pųipraven k experimentłm. Ukažme si nynķ napų. zjištģnķ, jak bude systém reagovat na plynulé zhoršovįnķ životnķho prostųedķ, které zpłsobķ, že bģhem nįsledujķcķch 100 let dojde ke snķženķ relativnķ rodivosti koųisti na polovinu současné hodnoty a současnģ zvżšenķ relativnķ śmrtnosti dravcł o polovinu současné hodnoty. Vżsledek simulace vidķme na obr. 4.33 {soubor KOR_DRA2.SIP}.
Zįvģr
Sestavili jsme zjednodušenż matematickż model systému koųist-dravec a provedli simulaci jeho chovįnķ. Srovnįnķm s namģųenżmi hodnotami byla zjištģna dobrį shoda se skutečnostķ. Pomocķ simulačnķch experimentł jsme pak mohli pųedpovģdģt jakż bude dalšķ vżvoj tohoto systému v zįvislosti na płsobenķ človģka.
Poslednķm modelem ( pųedchozķ modely byly systém pųķjmu potravy a systém koųist-dravec ) je zjednodušenż systém pųenosu energie v jezernķm ekosystému. Celż systém je pomģrnģ komplikovanż, jak vidķme na obr. 4.34.
Pro naše potųeby provedeme zjednodušenķ, budeme pųedpoklįdat, že systém obsahuje tųi prvky - rostliny, herbivory a karnivory. Vstupem do systému je energie slunečnķho zįųenķ. Stav jednotlivżch prvkł popisujķ stavové veličiny, které vyjadųujķ množstvķ biomasy udané jako energie vįzanį v biomase vztaženį na jednotku plochy nebo objemu. Na obr. 4.35 vidķme schéma pųedįvįnķ biomasy za jednotku času.
Čįst biomasy se pųitom ztrįcķ respiracķ a čįst jinżm propadem energie (odumķrįnķ, exkrementy). Konstanty uvedené v obr. 4.35 udįvajķ rychlost pųenosu energie z jednoho prvku systému do druhého, jejich rozmģrem je [rok-1]. Za jednotku času se tedy pųenese množstvķ energie dané součinem konstanty a aktuįlnķho množstvķ biomasy ve zdrojovém prvku.
Matematickż model pųenosu energie v jezernķm ekosystému
Celż systém pak młžeme popsat pomocķ trojice stavovżch rovnic (člen 1/365 sloužķ ke zmģnģ rozmģru konstant z [rok-1] na [den-1]) :
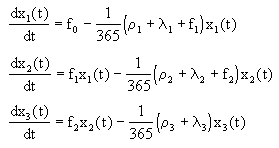
kde je
f0 - energetickż vstup slunečnķ energie [kJ m-2den-1],
x1 - energie biomasy rostlin [kJ m-2],
x2 - energie biomasy herbivorł [kJ m-2],
x3 - energie biomasy karnivorł [kJ m-2],
![]() i - rychlost ztrįty biomasy respiracķ [rok-1],
i - rychlost ztrįty biomasy respiracķ [rok-1],
![]() i - rychlost ostatnķch ztrįt biomasy [rok-1],
i - rychlost ostatnķch ztrįt biomasy [rok-1],
fi - rychlost toku biomasy mezi prvky systému.
Energetickż vstup slunečnķ energie do systému je pųitom popsįn rovnicķ :
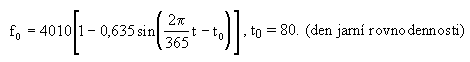
Prłbģh vstupu slunečnķ energie je zobrazen na obr. 4.36.
Hodnoty potųebnżch konstant uvįdķ nįsledujķcķ tabulka :
| 1 | rostliny | 250 000 | 0,48 | 1 | 2,55 |
| 2 | herbivoųi | 7 000 | 4,85 | 6,9 | 6,12 |
| 3 | karnivoųi | 8 000 | 2,7 | 1,95 |
Simulačnķ model pųenosu energie v jezernķm ekosystému
Na obr. 4.37 {soubor JEZ_EKOS.SIP} je uveden simulačnķ model systému pųenosu energie v jezernķm ekosystému. Na obr. 4.38 vidķme vżsledek chovįnķ systému. Pro lepšķ zobrazenķ bylo pro prłbģh x2 a x3 použito jiné mģųķtko - na pravé stranģ grafu.
DYKYJOVĮ, D. aj. 1989. Metody studia ekosystémł. Praha: Academia, 1989.
FARB, P. 1977. Ekologie. Praha: Mladį fronta, 1977.
KŲĶŽOVĮ, E. aj. 1992. Všeobecnį ekológia. Zvojen: TU, 1992.
KVASNIČKOVĮ, D. 1991. Zįklady ekologie. Praha: SPN, 1991.
LOSOS, B. aj. 1984. Ekologie živočichł. Praha: SPN, 1984.
LOSOS, B. aj. 1987. Zįklady obecné ekologie. Praha: SPN, 1987.
MADER, S. S. 1988. Inquiry into Life. Iowa: Dubuque, Wm.C.Brown Publishers, 1988.
Metodika biomonitoringu v CHKO a NP. Praha: MŽP ČR, 1992.
NEUMANNOVĮ, M. a FARANA, R. 1994. Ekologie. Prozatķmnķ učebnķ text. Ostrava: Katedra ochrany životnķho prostųedķ HGF VŠB, 1994.
NAVĮK, M., FĮBER, J. a KUFUDAKI, O. 1993. Neuronové sķtģ a informačnķ systémy živżch organismł. Praha: Grada, 1993
ODUM, E. P. 1977. Zįklady ekologie. Praha: Academia, 1977.
PAZOUREK, J. 1992. Simulace biologickżch systémł. Praha, Grada, 1992.
PIVNIČKA, K. 1988. Ekologie. Praha: SPN, 1988.
SLAVĶKOVĮ, J. 1986. Ekologie rostlin. Praha: SPN, 1986.
STARR, C. 1991. Biology - Concept and Aplicantion.Wadsworts: Publishing Company, 1991.
STILING, P. D. 1992. Introductory Ecology. Prentice Hall, Inc., A Simon & Schuster Company, Englewood Cliffs, New Jersey, 1992.
TöLGYESSY, J. a FARGĮŠOVĮ, A. 1986. Zįklady ekológie a toxikológie. Bratislava: STU, 1986.
![]()